
Download
Organ weight index = Wet organ weight g whole body weight g × 100 95 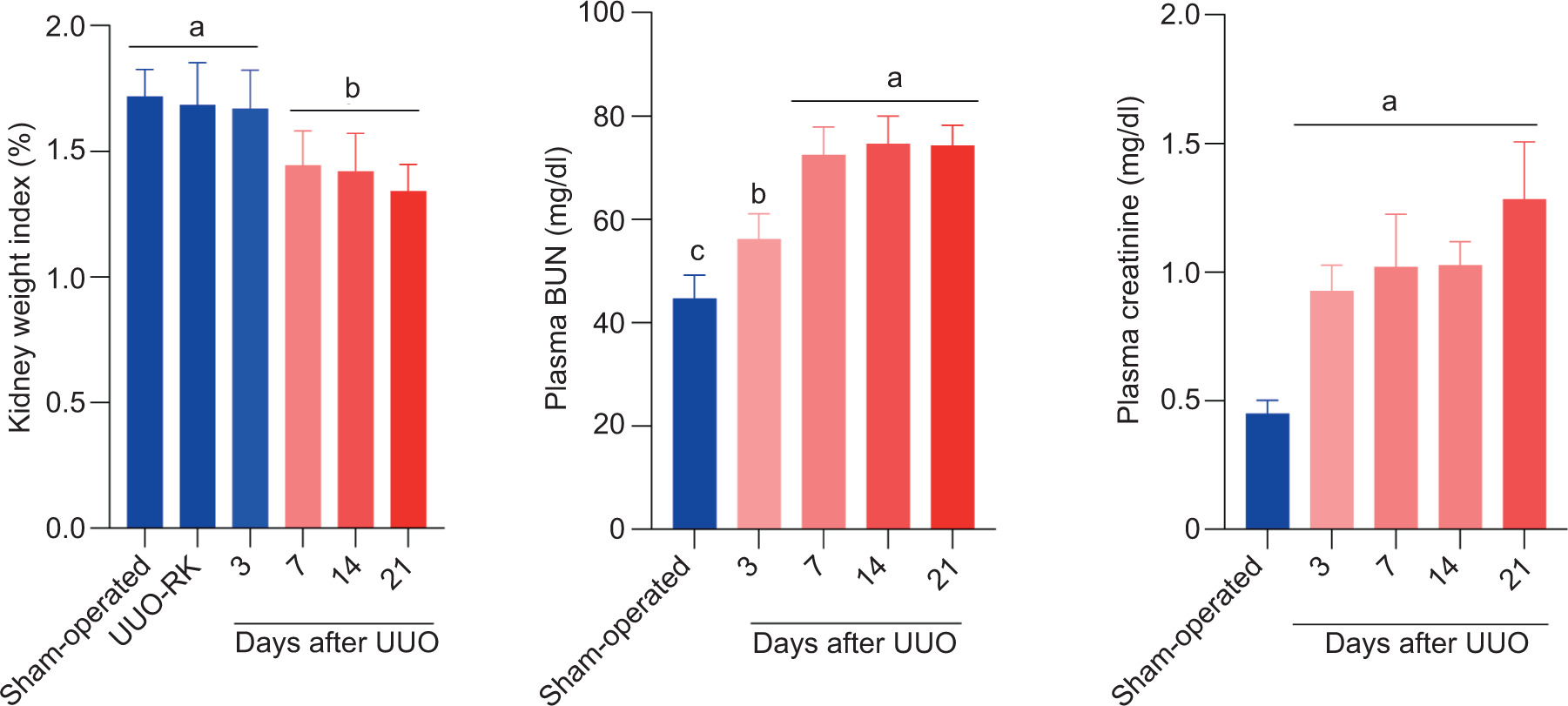
ORIGINAL ARTICLE: NEPHROLOGY
Mitochondrial Impairment and Oxidative Stress Are Essential Mechanisms Involved in the Pathogenesis of Acute Kidney Injury
Heresh Rezaei1,2, Zahra Honarpishefard1,2, Fatemeh Ghaderi1,2, Ayeh Rouhani1,2, Akram Jamshidzadeh1,2, Seyyed Mohammad Amin Kashani1,2, Narges Abdoli3, Forouzan Khodaei1,2, Omid Farshad1, Abdollah Arjmand4, Issa Sadeghian1,5, Negar Azarpira6, Mohammad Mehdi Ommati1,7,8*, Reza Heidari1
1Pharmaceutical Sciences Research Center, Shiraz University of Medical Sciences, Shiraz, Iran;
2Department of Pharmacology and Toxicology, School of Pharmacy, Shiraz University of Medical Sciences, Shiraz, Iran;
3Iran Food and Drug Administration, Ministry of Health and Medical Education, Tehran, Iran;
4Department of Pharmacology and Toxicology, Faculty of Pharmacy, Shahid Beheshti University of Medical Sciences, Tehran, Iran;
5Biotechnology Incubator, Shiraz University of Medical Sciences, Shiraz, Iran;
6Transplant Research Center, Shiraz University of Medical Sciences, Shiraz, Iran;
7College of Life Sciences, Shanxi Agricultural University, Taigu, Shanxi, China;
8Henan Key Laboratory of Environmental and Animal Product Safety, Henan University of Science and Technology, Luoyang, Henan, China
Abstract
Acute kidney injury (AKI) is an emergency condition that requires restrictive and appropriate clinical interventions. Identifying mechanisms of organ injury is a critical step in developing clinical interventions. Unilateral ureter obstruction (UUO) is widely used as an animal model for investigating AKI. The current study was designed to evaluate the role of mitochondrial impairment and oxidative stress in the pathogenesis of renal injury in UUO model. Mice underwent UUO surgery. Then, kidney tissue histopathological changes, plasma biomarkers of renal injury, oxidative stress, and different renal mitochondrial indices were evaluated at scheduled time intervals (3, 7, 14, and 21 days after UUO surgical procedure). Significant increase in plasma creatinine and blood urea nitrogen levels was evident in UUO mice. The UUO surgery induced severe kidney tissue histopathological alterations, including necrosis, severe tubular atrophy, and interstitial inflammation. Moreover, kidney biomarkers of oxidative stress included reactive oxygen species formation, lipid peroxidation, protein carbonylation, decreased glutathione reservoirs (GSH), and increased oxidized glutathione (GSSG) observed in UUO mice. On the other hand, significant mitochondrial depolarization, decreased mitochondrial dehydrogenases activity, mitochondrial permeabilization, and decreased adenosine triphosphate and GSH/GSSG levels were discovered in mitochondria isolated from the kidneys of UUO mice. The data obtained from the current study demonstrated a pivotal and interconnected role for oxidative stress and mitochondrial dysfunction in the pathogenesis of renal injury in UUO model. Therefore, these directions could serve as therapeutic targets in animal models or patients of acute renal failure.
Key words: antioxidants, bioenergetics, mitochondria, nephrotoxicity, renal failure
Received: 9 January 2023. Accepted after revision: 20 November 2023. Published: 26 December 2023
Authors for correspondence: Mohammad Mehdi Ommati, Henan Key Laboratory of Environmental and Animal Product Safety, Henan University of Science and Technology, Luoyang 471000, Henan, China. Email: mehdi.ommati@gmail.com; Reza Heidari, Pharmaceutical Sciences Research Center, Shiraz University of Medical Sciences, Shiraz, Iran, and PO Box 158371345, Roknabad Karafarin St. Shiraz, Fars, Iran. Email: rheidari@sums.ac.ir
How to cite: Rezaei H, et al. Mitochondrial Impairment and Oxidative Stress Are Essential Mechanisms Involved in the Pathogenesis of Acute Kidney Injury. J Ren Hepat Disord. 2023;7(2): 30–45.
Copyright: Rezaei H, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0
Introduction
Renal disease refers to a wide range of disorders ranging from mild kidney function abnormalities to failure of this vital organ. In this context, acute kidney injury (AKI) is an emergency clinical condition that needs appropriate and restricted clinical interventions to prevent renal failure (1, 2). A wide range of diseases and xenobiotics could cause AKI (1–6). The ultimate goal in patients with severe renal disease could be organ transplantation. However, detection of early tissue damage and understanding of the disease’s mechanisms could prevent/delay organ transplantation by developing appropriate therapeutic interventions.
The term oxidative stress refers to the situation where the balance between production and removal of reactive oxygen species (ROS) in biological environments is impaired (7). There is a plethora of evidence indicating the central role of oxidative stress and its associated complications in the pathogenesis of AKI (8–14). Increased levels of biomarkers of oxidative stress, including ROS formation, lipid peroxidation, oxidative damage of proteins, and other intracellular components (e.g., DNA), as well as a significant impairment of renal antioxidant defense mechanisms, have been documented in various models of AKI (15–19). Therefore, the role of oxidative stress and its association to mitochondrial impairment is highlighted in the current AKI model of unilateral ureter obstruction (UUO).
The association between oxidative stress and damage to vital organelles, such as mitochondria, is well documented (20–24). Mitochondria act as a powerhouse for producing cell energy. Meanwhile, mitochondria are also cells’ most critical ROS production center (20, 25–27). Therefore, there is a robust connection between oxidative stress and mitochondrial impairment.
Kidney tissue contains many mitochondria that, by producing sufficient energy, enable vital processes, such as reabsorbing substances in the kidney (27–30). Loss of mitochondrial function in renal tissue and consequent insufficient production of adenosine triphosphate (ATP) can lead to the urinary loss of vital electrolytes, vitamins, and minerals (29). Eventually, mitochondrial damage can lead to the release of cell death mediators from this organelle, leading to cytotoxicity, organ injury, and renal failure (29, 31, 32). The role of renal mitochondrial impairment and its association with oxidative stress are evaluated in the current model of UUO.
Obstructive renal injury and nephropathy are frequent clinical complications that could lead to AKI (33, 34). Despite their etiology, these complications could lead to irreversible consequences, such as renal failure. Therefore, identifying the mechanisms involved in the pathogenesis of obstructive kidney damage might help develop novel therapeutic strategies to control renal diseases. The current study aimed to evaluate the role of oxidative stress and mitochondrial impairment in the pathogenesis of renal injury in the UUO model of AKI. The data obtained from this study could help develop novel therapeutic strategies to combat AKI with different etiologies.
Materials and Methods
Reagents
2',7'-Dichlorofluorescein diacetate (DCFH-DA), reduced glutathione (GSH), trichloroacetic acid (TCA), malondialdehyde (MDA), 3-[4,5dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT), sucrose,3-(N-morpholino) propanesulfonic acid (MOPS), D-mannitol, ferric chloride hexahydrate (FeCl3.6H2O), rhodamine123 (Rh 123), thiobarbituric acid (TBA), 2,4,6-tripyridyl-s-triazine (TPTZ), dithiothreitol (DTT), coomassie brilliant blue, 2,4-dinitrophenyl hydrazine (DNPH), and 6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox) were purchased from Sigma (Sigma-Aldrich, USA). Kits for measuring serum biomarkers of renal injury were obtained from Pars-Azmun® (Tehran, Iran). Meta-phosphoric acid, n-propanol, ethylenediaminetetraacetic acid (EDTA), perchloric acid, and 2-amino-2-hydroxymethyl-propane-1, 3-diol-hydrochloride (Tris-HCl) were purchased from Merck (Darmstadt, Germany).
Animals
Male BALB/c mice (n = 60, 25±3 g) were obtained from Shiraz University of Medical Sciences, Shiraz, Iran. Animals were kept at an environmental temperature of 23±1ºC with ≈40% relative humidity and adequate ventilation. Animals had free access to tap water and a standard pellet chow during experiments. The Institutional Laboratory Animal Care and Use Committee at Shiraz University of Medical Sciences Shiraz, Iran, approved all animal experiments (Code: IR.SUMS.REC.1398.1220). The Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines for the care and use of experimental animals were also followed.
Unilateral ureter obstruction model of acute kidney injury
Animals were randomly divided into the UUO and sham-operated groups. The UUO model was induced based on the previously reported protocol (35). Briefly, mice were anesthetized with a mixture of 8 mg/kg xylazine and 60 mg/kg ketamine. The left ureter was isolated and doubly ligated. The sham-operated mice underwent an identical surgical intervention for ureter identification and manipulation without ligation (35). Animals were recovered under Infrared (IR) light and received normal saline (2.5 mL/kg subcutaneous [s.c.]). Mice had access to easy food on the first day after surgery (36). No mortality rate was discovered in the current study.
Experimental setup and sample collection
Animals were intensely anesthetized with thiopental, 100 mg/kg, at different time intervals (3, 7, 14, and 21 days after UUO procedure). Blood samples (1 mL from the inferior vena cava) were transferred to sodium citrate-coated tubes, and the plasma was prepared (4000 g, 20 min, 8°C). Afterward, five mice in each group were randomly selected for kidney mitochondria isolation. Kidneys of five other mice were used to assess histopathological alterations and oxidative stress biomarkers. All mentioned markers were also evaluated in the right intact kidney (without ureter obstruction; as a control in each animal) on day 21 post-UUO surgery of right kidney (UUO-RK).
Reactive oxygen species formation
Reactive oxygen species were estimated in the kidneys of UUO mice using the 2',7'-dichlorofluorescein diacetate (DCF-DA) method (9, 37–41). Briefly, 200 mg of tissue samples were homogenized in 5 mL of ice-cooled Tris-HCl buffer (40 mM, pH = 7.4). Then, 100 µL of the resulting tissue homogenate was mixed with 900 µL of Tris-HCl buffer (40 mM, pH = 7.4) and 10 µL of DCF-DA (final concentration of 10 µM) (3, 42–47). Samples were incubated in dark for 10 min (37ºC shaker incubator). Finally, the fluorescence intensity was assessed using a FLUOstar Omega® fluorimeter (λexcit = 485 nm and λemiss = 525 nm) (9, 37, 40, 48, 49).
Protein carbonylation
Renal tissue protein carbonylation in UUO mice was assessed based on the dinitrophenyl hydrazine (DNPH) test (50–52). Briefly, kidney tissue (200 mg) was homogenized in 5-mL phosphate buffer (pH = 7.5, containing 0.1% v:v of triton X-100). Samples were centrifuged (700 ×g, 10 min, 4ºC) and the resulting supernatant was treated with 1500 µL of 10-mM DNPH solution (dissolved in 6-M HCl). Samples were incubated in a shaker incubator (for 1 h, 25ºC, protected from light) (50, 51, 53–56). Afterward, 500-µL trichloroacetic acid (20% w:v) was added and centrifuged (17,000 ×g, 5 min, 4ºC). The pellet was washed for five times with ethanol:ethyl acetate (1 mL of 1:1 v:v) and redissolved in guanidine chloride (6 M, pH = 2.3). Finally, samples were centrifuged (17,000 ×g, 1 min, 4ºC), and the absorbance of the supernatant was assessed (λ = 370 nm) (51, 57).
Lipid peroxidation
The thiobarbituric acid reactive substances (TBARS) test was used to assess lipid peroxidation in the kidneys of UUO mice (9, 47, 58–62). For this purpose, 0.5 mL of tissue homogenate (10% w:v in Tris-HCl buffer, 40 mM, pH = 7.4) was treated with 1 mL of TBARS assay reagent (a mixture of 0.4% w:v of thiobarbituric acid, 50% w:v of trichloroacetic acid, and 1% w:v of meta-phosphoric acid, pH = 2) (9, 42, 63–70). Samples were vortexed (1 min) and heated (100 C water bath) for 45 min. Then, 1 mL of n-butanol was added. Samples were mixed considerably and centrifuged (10,000 ×g, 20 min, 25°C). Finally, absorbance of the upper phase was measured (λ = 532 nm) (9, 71–74).
Renal hydroxyproline levels
Renal hydroxyproline content was assessed as an index of tissue fibrosis. Briefly, 500 µL of tissue homogenate (10% w: v in Tris-HCl buffer) was digested in 1 mL of hydrochloric acid (6 N) at 120°C (12 h). Afterward, an aliquot of digested homogenate (250 μL) was treated with 250 μL of citrate-acetate buffer (pH = 6) and 500 μL of chloramines-t (56 mM) and incubated at room temperature for 20 min. Then, 500 μL of Ehrlich’s reagent (15 g of p-dimethyl amino benzaldehyde in n-propanol/perchloric acid; 2:1 v:v) was added and incubated at 65°C (15 min). Finally, the absorbance of the developed color was measured at λ = 550 nm (75).
Total antioxidant capacity of the kidney tissue
The ferric-reducing antioxidant power (FRAP) assay measured the total antioxidant capacity of renal tissue (9, 76). Tissue was homogenized in an ice-cooled (4ºC) 40-mM Tris-HCl buffer. Afterward, 100 µL of tissue homogenate (10% w:v in Tris-HCl buffer) was added to 900 µL of freshly prepared FRAP reagent (77–82). Samples were incubated at 37ºC (5 min, protected from light). Finally, the absorbance was measured at λ = 595 nm (9, 83).
Myeloperoxidase enzyme activity in the kidney
Myeloperoxidase (MPO) activity of the renal tissues of UUO mice was assessed based on the previously reported procedure (84). Briefly, tissue specimens (100 mg) were homogenized in 1 mL of hexadecyl-trimethyl-ammonium bromide (HTAB) solution (0.5% w:v of HTAB; dissolved in 50-mM potassium phosphate buffer; pH = 6, at 4°C) and centrifuged (3000 ×g, 20 min, at 4°C). Then, 100 µL of the supernatant was added to 2.9 mL of 50-mM potassium phosphate buffer (pH = 6; containing 16.7-mg/100 mL of O-dianisidine hydrochloride and 0.0005% v:v of H2O2). Samples were incubated in dark for 5 min (25°C). Then the reaction was stopped by HCl (100 µL of 1.2 M). Finally, the absorbance was measured at λ = 400 nm (84).
Kidney mitochondria isolation
The differential centrifugation method was used to isolate kidney mitochondria (85–89). For this purpose, mice kidneys were washed and minced in an ice-cold buffer medium (220-mM sucrose, 2-mM HEPES, 0.5-mM EGTA, 70-mM mannitol, and 0.1% bovine serum albumin (BSA), pH = 7.4). Then, the minced tissue was transported into a fresh buffer (5-mL buffer/1 g of the kidney) and homogenized. The homogenized tissue was centrifuged (1000 ×g, 20 min, 4°C), and the supernatant was collected (90–93). The supernatant was centrifuged again (10,000 ×g, 20 min, 4°C) to pellet mitochondrial fraction. The mitochondrial pellet was washed for at least three times using fresh isolation buffer to increase mitochondrial purity and yield. Finally, isolated mitochondria were suspended in buffer and used for further evaluation.
Mitochondrial dehydrogenases activity
The 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay was applied to determine renal mitochondrial dehydrogenases activity (8, 44, 94–96). Briefly, a mitochondrial suspension (0.5-mg protein/mL) was incubated with 40 µL of 0.4% w:v of MTT solution (37°C, 30 min, protected from light) (85, 97). Then the samples were centrifuged (10,000 ×g, 20 min), and the pellet was dissolved in dimethyl sulfoxide (DMSO, 1000 µL). Finally, the absorbance was measured at λ = 570 nm (98–100).
Mitochondrial depolarization
Briefly, mitochondrial fractions (0.5-mg protein/mL) were incubated with 10 µM of Rh 123 at 37ºC in a shaker incubator (10 min, protected from light) (101–104). Afterward, samples were centrifuged (10,000 ×g, 10 min, 4ºC), and the fluorescence intensity of the supernatant was assessed (FLUOstar Omega® Fluorimeter; λexcitation = 485 nm and λemission = 525 nm) (9, 105–107).
Mitochondrial swelling
Analysis of mitochondrial swelling was estimated through changes in light scattering (105, 108–114). Briefly, isolated mitochondria samples (0.5-mg protein/mL) were added to pre-warmed buffer. The absorbance was monitored at λ = 540 nm for 30 min at 30°C with orbital shaking (EPOCH® microplate reader; Agilent Technologies, Santa Clara, CA, USA). Primary and final absorbance difference (ΔOD) was calculated (28, 105, 115).
Renal tissue and isolated mitochondrial glutathione content
The reduced (GSH) and oxidized (GSSG) glutathione content in the kidneys of UUO mice was measured using the high-performance liquid chromatography (HPLC) method based on derivatization with dinitrofluorobenzene (DNFB) (3, 116, 117). An amine column (NH2, 25-cm Bischoff chromatography; Leonberg, Germany) was used as a stationary phase (118). Buffer A (acetate buffer:water; 1:4 v/v) and buffer B (methanol:water; 4:1 v/v) were mobile phases. Gradient method with a regular increase of buffer B to 95% in 30 min was used, and the flow rate was 1 mL/min (119). The ultraviolet (UV) detector was set at λ = 254 nm.
Mitochondrial ATP content
The previously described HPLC protocol assessed mitochondrial ATP level (120, 121). Briefly, isolated mitochondria (0.5-mg protein/mL) were treated with ice-cooled 0.2-M perchloric acid, incubated in ice (5 min) and centrifuged (10 min, 17,000 ×g, 4ºC). The supernatant was treated with an equivalent volume of ice-cooled 1-M KOH solution. Samples were filtered and injected (30 µL) into an HPLC system consisting of an LC-18 column (µ-Bondapak, 15 cm). The mobile phase comprised KH2PO4 (215 mM), tertiary butyl ammonium sulfate (2.3 mM), KOH (1 M, 0.4%), and acetonitrile (4% v:v). The constant flow rate was 1 mL/min, and the UV detector was set at λ = 254 nm (5).
Tissue histopathology and organ weight index
Kidney samples were fixed in buffered formalin (10% w:v formaldehyde in 0.1-mM phosphate buffer, pH = 7.4). Then, paraffin-embedded kidney sections (5 µm) were prepared and stained with hematoxylin and eosin (H&E) (122). Renal histopathological changes in UUO model were scored based on the previously reported following protocol: score 0, none; score 0.5, <10%; score 1, 10–25%; score 2, 25–50%; score 3, 50–75%, and score 4, >75% (123). Masson’s trichrome staining determined fibrotic kidney changes in UUO (21). Periodic acid–Schiff (PAS) staining was applied to assess kidney cast formation. A pathologist analyzed samples blindly. Kidney weight index was measured as follows:
Statistical analysis
Data were presented as mean ± SD. Data comparison was accomplished by one-way analysis of variance (ANOVA) with post hoc Tukey’s multiple comparison test. The normality of data sets was assessed by Kolmogorov–Smirnov test. Scores for histopathological changes in renal tissue were presented as median and quartiles for five random scores. The analysis of tissue histopathological alterations was performed by the Kruskal–Wallis test, followed by the Mann–Whitney U test. P < 0.05 was considered significant.
Results
Kidney weight index was significantly decreased in UUO mice after 7, 14, and 21 days of surgery (Figure 1). No significant changes were recorded in the right kidney of UUO mice (UUO-RK), compared to the sham-operated group (Figure 1).
Figure 1: Serum biochemical measurements and renal weight index in unilateral ureter obstruction (UUO) model of acute kidney injury. UUO-RK: Right kidney of UUO model. Data are presented as mean ± SD (n = 5). Groups with different alphabetical superscripts are significantly different (P < 0.05).
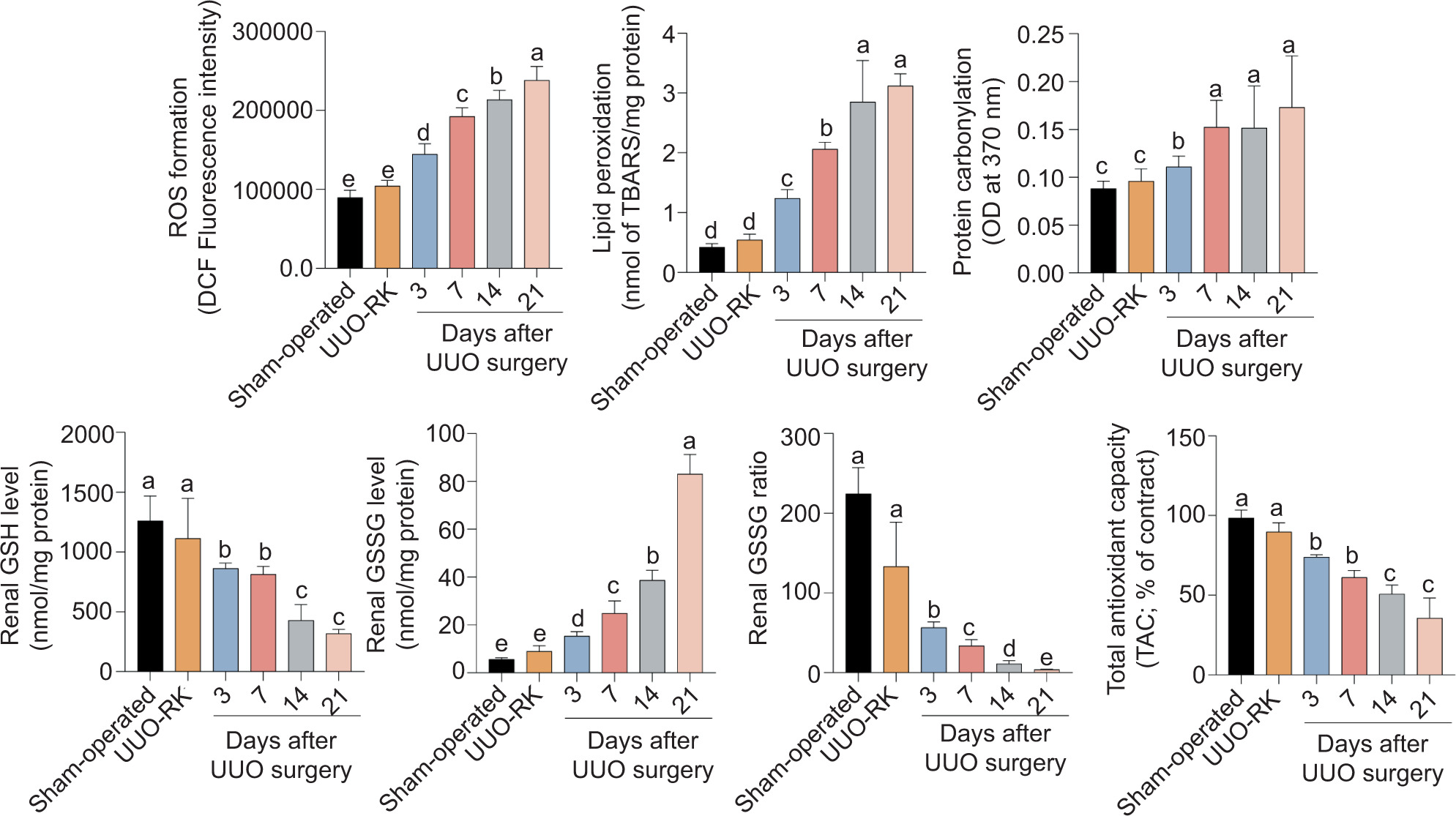
Plasma blood urea nitrogen (BUN) and creatinine levels as biomarkers of renal injury were significantly elevated at different time points of post-ureteral obstruction (Figure 1). On the other hand, oxidative stress markers were altered in both sham-operated and UUO model (Figure 2). A time-dependent increase in renal tissue ROS, lipid peroxidation, protein carbonylation, and increased GSSG levels was evident in UUO mice (Figure 2). Moreover, renal antioxidant capacity reduced GSH content and GSH–GSSG ratio was significantly decreased at different time intervals after UUO surgical procedure in a time-dependent manner (Figure 2). No significant changes in renal biomarkers of oxidative stress were recorded in the right kidney of UUO mice (UUO-RK), compared to the sham-operated group (Figure 2).
Figure 2: Biomarkers of oxidative stress in the renal tissue of unilateral ureter obstruction (UUO) mice model of acute kidney injury. UUO-RK: Right kidney of UUO model. Data are presented as mean ± SD (n = 5). Groups with different alphabetical superscripts are significantly different (P < 0.05).
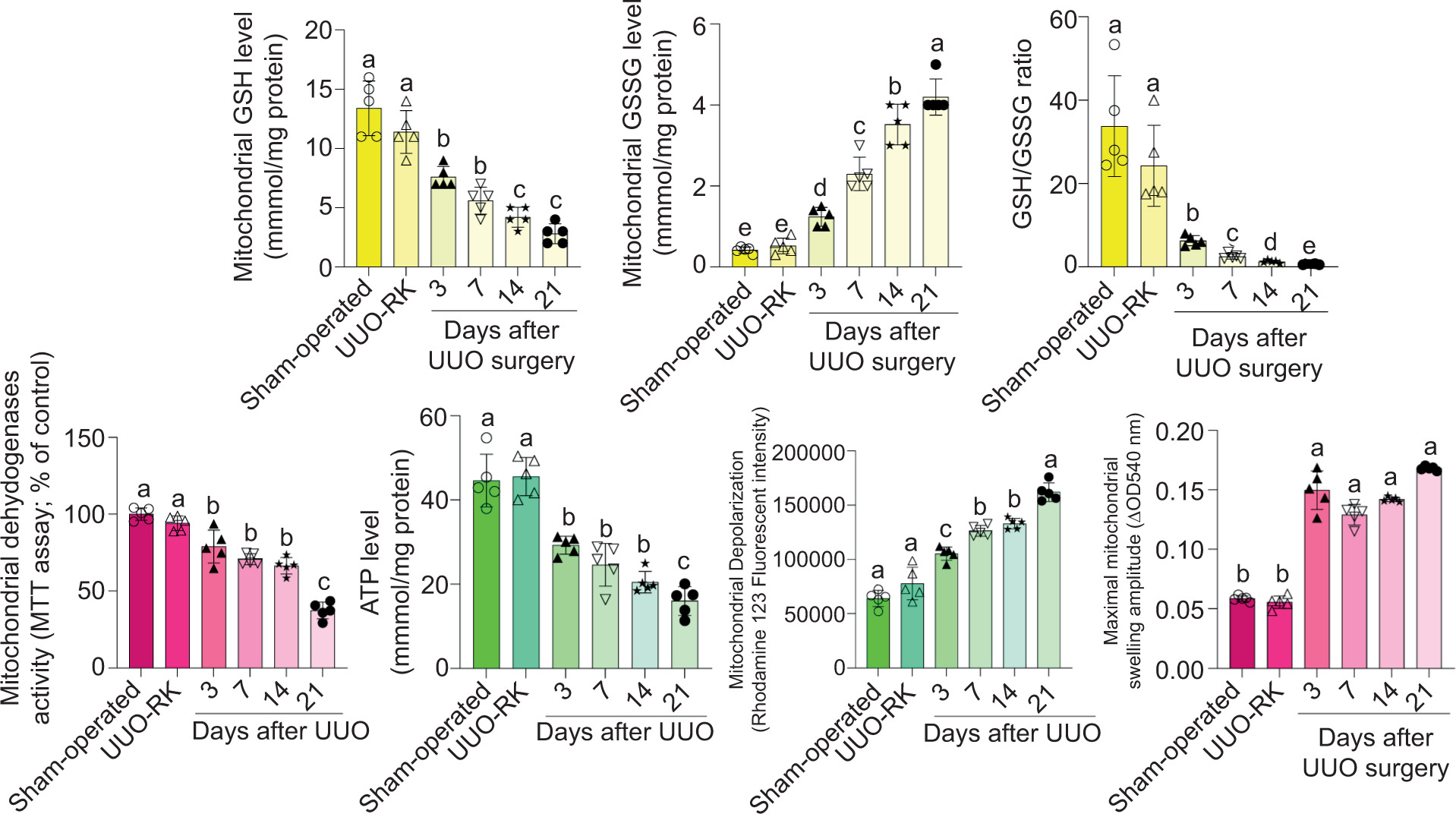
A significant decrease in mitochondrial dehydrogenase activity and ATP levels was evident in the kidney of UUO model (Figure 3). On the other hand, severe mitochondrial permeabilization and depolarization were also detected in mitochondria isolated from UUO mice in a time-dependent manner (Figure 3). The GSH reservoirs were also depleted, and the GSH–GSSG ratio decreased considerably in the mitochondria isolated from UUO mice (Figure 3). No significant changes in mitochondrial indices were discovered when the right kidney of UUO mice (UUO-RK) was compared with that of the sham-operated group (Figure 3).
Figure 3: Mitochondrial indices in the kidney of unilateral ureter obstruction (UUO) mice. UUO-RK: Right kidney of UUO model. Data are presented as mean ± SD (n = 5). Groups with different alphabetical superscripts are significantly different (P < 0.05).
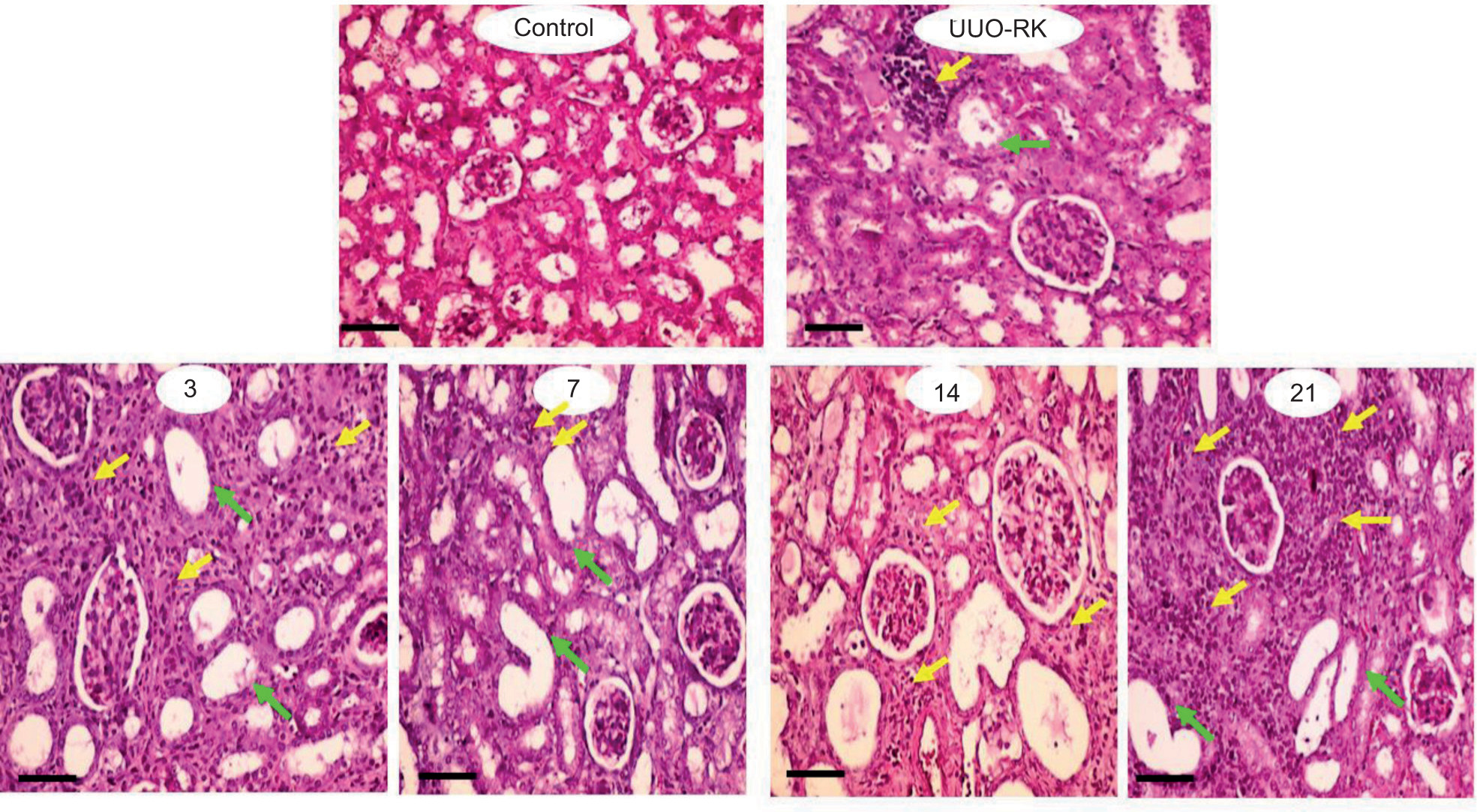
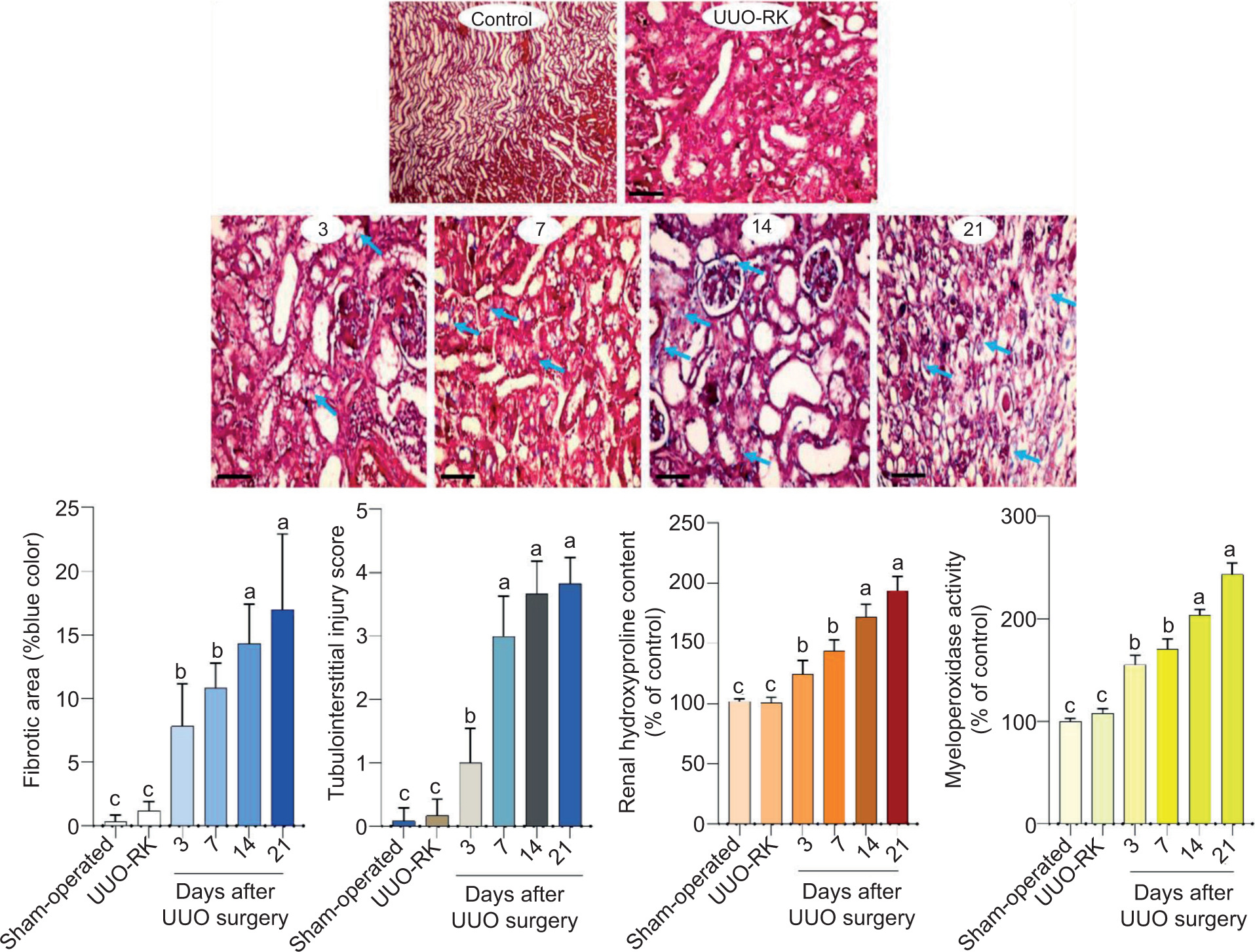
Renal tissue histopathological assessments revealed significant interstitial inflammation, tubular atrophy, and tissue necrosis on different days post-UUO surgery (Figure 4 and Table 1). A mild inflammatory response was also apparent only in the right kidney of UUO mice (UUO-RK) on day 21 after UUO surgery (Table 1). Markers of kidney fibrosis were also evaluated in both sham-operated and UUO mice (Figure 5). Collagen deposition was significantly elevated in the kidneys of UUO mice than in the kidneys of sham-operated mice (Figure 5). Renal tissue hydroxyproline content was also considerably increased in UUO mice (Figure 5). MPO enzyme activity as an index of tissue inflammation was also significantly higher than in the control animals at various time intervals post-UUO surgery (Figure 5).
Figure 4: Renal histopathological changes in unilateral ureter obstructed (UUO) mice. Numbers (3, 7, 14, 21) indicate days after the UUO surgery. Inflammatory cell infiltration (yellow arrows) and tubular atrophy (green arrows) were prominent histopathological changes in UUO mice. UUO-RK: Right kidney of UUO model. Scale bar = 100 µm.
Figure 5: Renal fibrotic changes, collagen deposition (blue arrow), and hydroxyproline content in the unilateral ureter obstruction (UUO) model (Trichrome stain ×200). Numbers on histopathological pictures (3, 7, 14, 21) indicate days after UUO surgery. UUO-RK: Right kidney of UUO model. Data for fibrotic area, hydroxyproline content, myeloperoxidase activity, and tubulointerstitial injury score are given as mean ± SD (n = 5). Columns with various alphabetical superscripts are significantly different (P < 0.05). Scale bar = 100 µm.
Table 1: Renal histopathological alterations in unilateral ureter obstruction (UUO) mice.
| Treatments | Glomerular Damage | Tubular Damage | Inflammation |
|---|---|---|---|
| Control | 0 (0, 0)a | 0 (0, 0)a | 0 (0, 0)a |
| UU-RK | 0 (0, 0)a | 0 (0, 0)a | 0.5 (0, 1)a |
| 3 days after UUO | 2 (1, 2)b | 2 (2, 2)b | 3 (3, 3)d |
| 7 days after UUO | 3 (2, 3)d | 3 (3, 3)d | 4 (4, 4)e |
| 14 days after UUO | 3 (3, 3)d | 4 (4, 4)e | 4 (4, 4)e |
| 21 days after UUO | 4 (4, 4)e | 4 (4, 4)e | 4 (4, 4)e |
Data are presented as median and quartiles for five pictures/groups.
Data sets with different alphabetical superscripts are significantly different (P < 0.05).
Renal histopathological changes in UUO model were scored based on a protocol described by Li et al. (2019) in the materials and methods section (123). UU-RK: Right kidney of UUO mice.
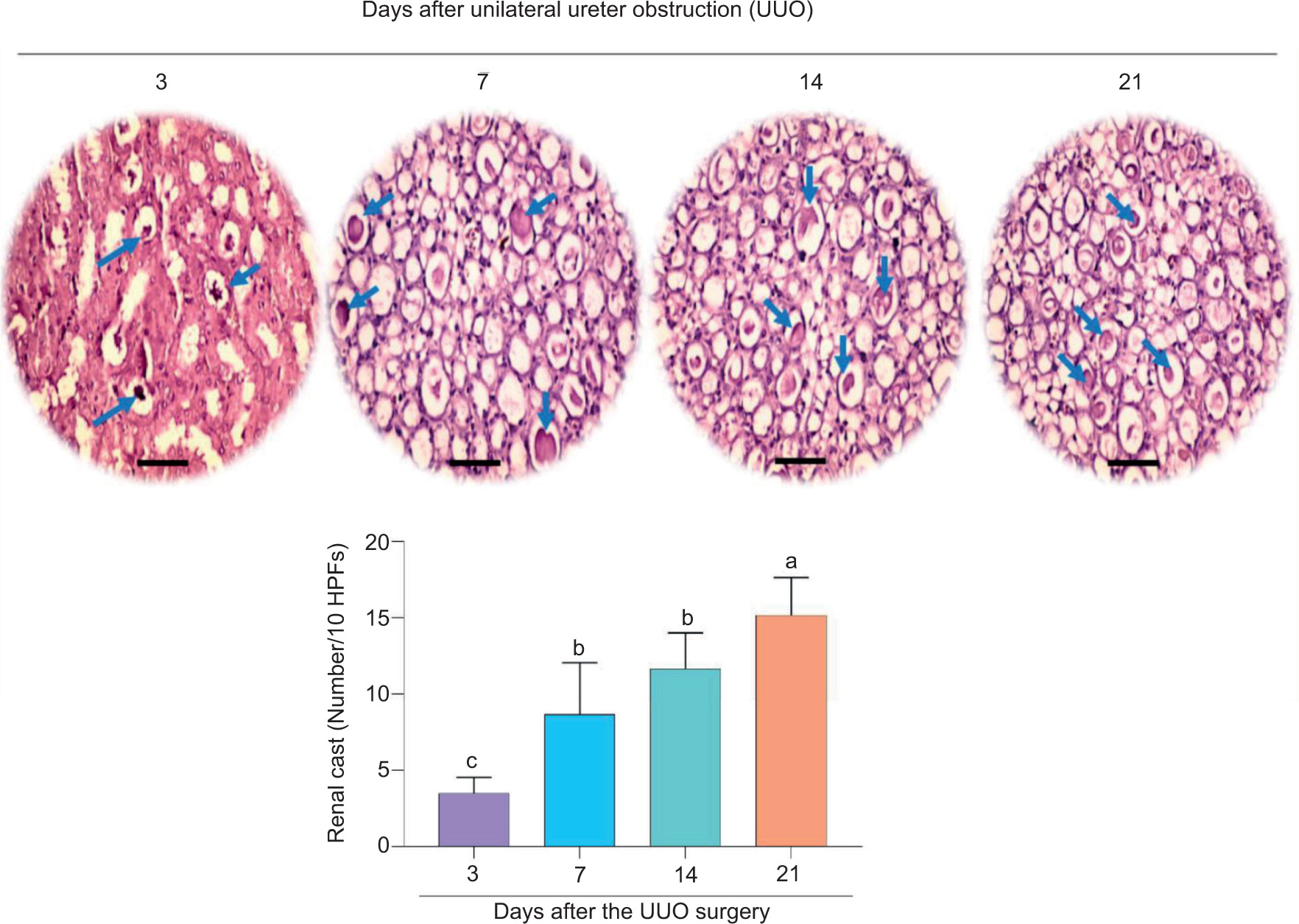
Periodic acid–Schiff staining of renal tissue revealed significant cast formation in UUO mice (Figure 6). It was discovered that the number of kidney casts was time-dependently increased in the kidney after UUO surgery (Figure 6).
Figure 6: Cast formation in the kidney of unilateral ureter obstruction (UUO) model (periodic acid–Schiff [PAS] stain ×400). No significant cast formation was detected in the sham-operated group or right kidney of UUO model. HPFs: high power fields. Data for renal cast formation are presented as mean ± SD (n = 5). Columns with various alphabetical superscripts are significantly different (P < 0.05). Scale bar = 100 µm.
Discussion
Acute kidney injury is a severe clinical complication requiring restricted and emergent interventions (1, 2). Determination of the mechanisms involved in the pathogenesis of the diseases is a critical step in identifying and developing therapeutic targets. UUO is a model investigated widely for evaluating the modality of renal injury. Inflammatory response, mechanical stress, and oxidative stress are commonly assessed in the UUO model of renal injury. On the other hand, there is no report on the role of mitochondrial impairment in cell death and organ injury in UUO model. In the current study, we discovered that severe oxidative stress, mitochondrial impairment, inflammation, and renal tissue histopathological alterations occurred in the kidney of UUO model. These findings demonstrated essential role of mitochondrial function disturbances and oxidative stress in the pathogenesis of renal injury in UUO model.
As mentioned, identifying the cellular and molecular mechanisms involved in the pathogenesis of diseases is a critical step in developing therapeutic strategies. In this context, complications, such as prostatic hyperplasia and renal stones, could lead to urinary obstruction (124, 125). Although surgical intervention is an ultimate and promising therapeutic option for this condition, finding organ injury mechanisms could help develop pharmacological or ancillary treatments.
In the current study, we investigated the role of oxidative stress and mitochondrial impairment and their interconnection in the pathogenesis of AKI in an animal model. Previous evidence indicated the role of oxidative stress and its associated complications in UUO model (126–128). It was discovered that oxidative stress markers, such as ROS formation and damage to cellular components (e.g., lipids), occurred in UUO model (126–128). Moreover, it was discovered that antioxidant mechanisms in renal tissues were significantly impaired in UUO model (126, 127). However, no specific source(s) for ROS was identified in UUO model yet. In the current study, we determined that oxidative stress biomarkers were significantly elevated in the kidneys of UUO mice (Figure 3). Moreover, we tried to delineate a connection between oxidative stress and potential sources of ROS formation in UUO model. In this regard, we investigated the relation between inflammatory response, mitochondrial impairment, and oxidative stress in the UUO model of AKI.
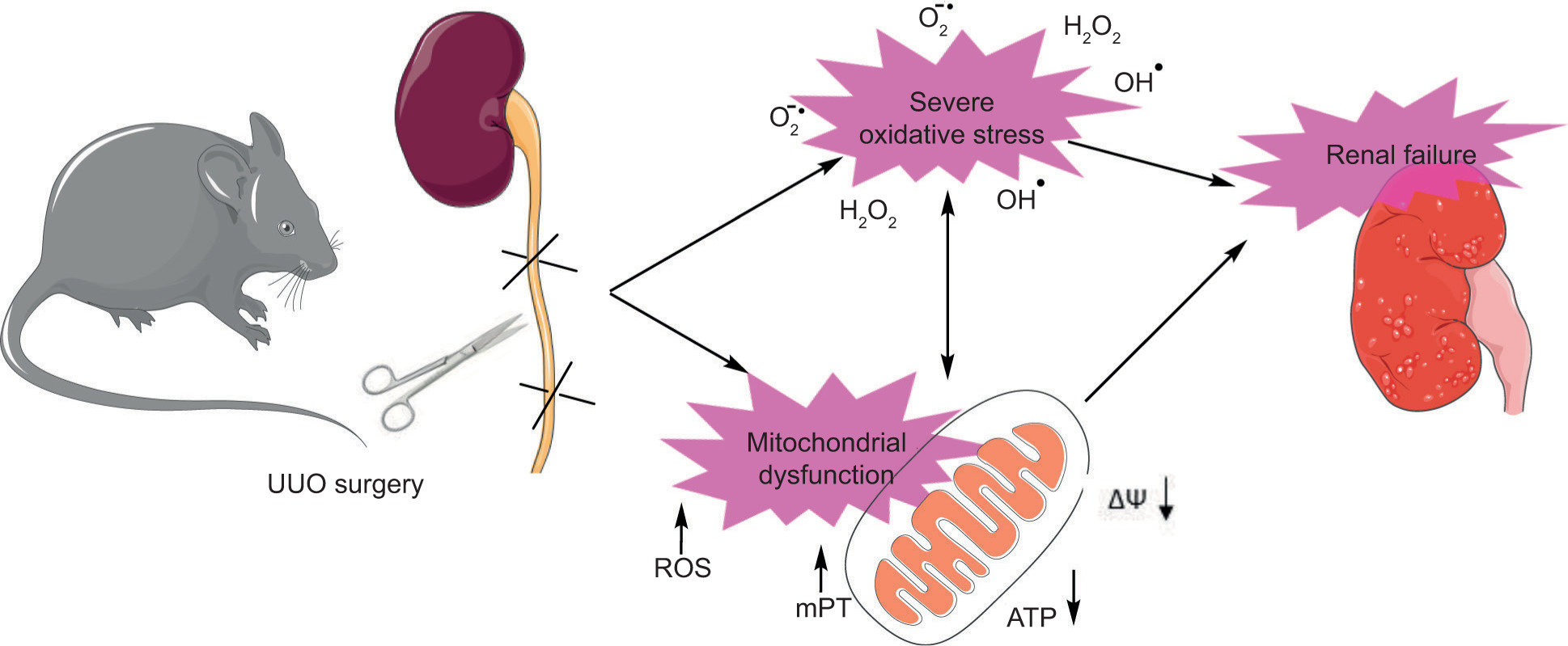
An interesting point was the interconnection between mitochondrial impairment and oxidative stress (Figure 7). It is well known that mitochondria are the primary source of intracellular ROS formation. An average level of mitochondrial-mediated ROS formation could act as signaling molecules essential for normal physiological processes (20). However, the mitochondrial-mediated ROS level increased dramatically if the mitochondrial function was impaired (20, 101). In the current UUO model, accumulating potently cytotoxic molecules routinely excreted from urine could affect mitochondrial function and lead to cytotoxicity and organ injury.
Figure 7: Schematic presentation of the central role of oxidative stress and mitochondrial impairment in the pathogenesis of unilateral ureter obstruction (UUO) model of acute renal injury. Oxidative stress and mitochondrial injury are two mechanistically interrelated events. ROS: reactive oxygen species; mPT: mitochondrial permeability transition; ATP: adenosine triphosphate; ΔΨ: mitochondrial membrane potential.
Previous studies revealed that inflammatory response is essential for renal injury in UUO model (128, 129). In the current study, we determined that the accumulation of inflammatory cells increased significantly at different time intervals after UUO surgery (Figure 4 and Table 1). The cytokines released by these cells could substantially cause oxidative stress in this model. On the other hand, it was discovered that cytokines, such as tumor necrosis factor-α (TNF-α), directly affected mitochondrial function (130, 131). These cytokines were also able to induce tissue fibrosis, finally leading to organ failure. It should also be mentioned that oxidative stress and inflammation are two interrelated processes that could lead to tissue damage (132, 133). The molecular connection between oxidative stress and inflammation is complicated and includes several signaling pathways (132, 133). The transcription factors nuclear factor erythroid 2-related factor 2 (Nrf-2) and nuclear factor kappa B (NF-κB) are critical oxidative stress and inflammation regulators, respectively (3, 6). Hence, more studies on these molecular pathways could provide viable therapeutic options to combat renal injury (e.g., in UUO model). Therefore, further research is needed to uncover this hypothesis.
Novel therapeutic options, such as mitochondrial replacement therapy, could be crucial in the future therapeutic strategies to control renal failure with different etiologies. These organelles provide enough energy for cell survival, prevent electrolyte imbalance, and provide enough time for basic organ transplantation strategies.
It has been well-documented that oxidative stress and the subsequent activation of extracellular matrix-producing cells (collagen deposition) are interconnected. Hence, a possible mechanism for fibrotic change in the current UUO model could be associated with oxidative stress (e.g., mitochondrial-mediated ROS formation). Kidney tissue also contains numerous mitochondria, guaranteeing the reabsorption of essential chemicals from urine (29). This process is an energy-dependent reaction. Hence, changes in renal mitochondrial function could lead to crucial electrolyte waste and serum electrolyte imbalance (8, 134, 135). Therefore, prior to surgical interventions, it is vital to preserve mitochondrial function and prevent kidney injury. Cellular mitochondria play a critical role in cell death (136).
Various studies revealed the significance of oxidative stress in UUO model (126, 127). Hence, our data on the role of oxidative stress markers are in line with these studies (Figure 7). More importantly, we found that renal mitochondrial indices of functionality were significantly hampered in UUO model. This represents mitochondrial function as a key mechanism in the pathogenesis of this complication (Figure 7). Recently, we tested different safe and clinically applicable agents with antioxidant and mitochondrial protecting agents in various experimental models or patients with renal disorders (5, 42, 91, 137–139). Some of these compounds even underwent clinical trials for different types of renal injury and nephropathy. Further studies are warranted to reveal the clinical significance of these data in complications leading to ureter obstruction and renal injury (e.g., prostate hyperplasia or kidney stones).
Acknowledgments
The authors acknowledge the Pharmaceutical Sciences Research Center for providing technical facilities for this investigation. This study was financially supported by the vice chancellor of research affairs of Shiraz University of Medical Sciences (Grants #17231/23701/23031/23040/23028/16428) and the Natural Science Foundation of Shanxi (Grant #20210302124411).
Conflict of interest
The authors declared that they had no conflict of interest to disclose.
REFERENCES
1. Gameiro J, Fonseca JA, Outerelo C, Lopes JA. Acute kidney injury: From diagnosis to prevention and treatment strategies. J Clin Med. 2020;9(6):1704. 10.3390/jcm9061704
2. Kinsey GR, Okusa MD. Pathogenesis of acute kidney injury: Foundation for clinical practice. Am J Kidney Dis. 2011;58(2):291–301. 10.1053/j.ajkd.2011.02.385
3. Ahmadi A, Niknahad H, Li H, Mobasheri A, Manthari RK, Azarpira N, et al. The inhibition of NFκB signaling and inflammatory response as a strategy for blunting bile acid-induced hepatic and renal toxicity. Toxicol Lett. 2021;349:12–29. 10.1016/j.toxlet.2021.05.012
4. Ommati MM, Attari H, Siavashpour A, Shafaghat M, Azarpira N, Ghaffari H, et al. Mitigation of cholestasis-associated hepatic and renal injury by edaravone treatment: Evaluation of its effects on oxidative stress and mitochondrial function. Liver Res. 2021;5(3):181–93. 10.1016/j.livres.2020.10.003
5. Ommati MM, Niknahad H, Farshad O, Azarpira N, Heidari R. In vitro and in vivo evidence on the role of mitochondrial impairment as a mechanism of lithium-induced nephrotoxicity. Biol Trace Elem Res. 2021;199(5):1908–18. 10.1007/s12011-020-02302-9.
6. Mousavi K, Niknahad H, Li H, Jia Z, Manthari RK, Zhao Y, et al. The activation of nuclear factor-E2-related factor 2 (Nrf2)/heme oxygenase-1 (HO-1) signaling blunts cholestasis-induced liver and kidney injury. Toxicol Res. 2021;10(4):911–27. 10.1093/toxres/tfab073
7. Poprac P, Jomova K, Simunkova M, Kollar V, Rhodes CJ, Valko M. Targeting free radicals in oxidative stress-related human diseases. Trends Pharmacol Sci. 2017;38(7):592–607. 10.1016/j.tips.2017.04.005
8. Heidari R, Ahmadi A, Mohammadi H, Ommati MM, Azarpira N, Niknahad H. Mitochondrial dysfunction and oxidative stress are involved in the mechanism of methotrexate-induced renal injury and electrolytes imbalance. Biomed Pharmacother. 2018;107:834–40. 10.1016/j.biopha.2018.08.050
9. Shafiekhani M, Ommati MM, Azarpira N, Heidari R, Salarian AA. Glycine supplementation mitigates lead-induced renal injury in mice. J Exp Pharmacol. 2019;11:15–22. 10.2147/JEP.S190846
10. Ghanbarinejad V, Jamshidzadeh A, Khalvati B, Farshad O, Li H, Shi X, et al. Apoptosis-inducing factor plays a role in the pathogenesis of hepatic and renal injury during cholestasis. Naunyn Schmiedeberg Arch Pharmacol. 2021;394(6):1191–203. 10.1007/s00210-020-02041-7
11. Tomsa AM, Alexa AL, Junie ML, Rachisan AL, Ciumarnean L. Oxidative stress as a potential target in acute kidney injury. Peer J. 2019;7:e8046. 10.7717/peerj.8046
12. Sureshbabu A, Ryter SW, Choi ME. Oxidative stress and autophagy: Crucial modulators of kidney injury. Redox Biol. 2015;4:208–14. 10.1016/j.redox.2015.01.001
13. Ommati MM, Farshad O, Azarpira N, Ghazanfari E, Niknahad H, Heidari R. Silymarin mitigates bile duct obstruction-induced cholemic nephropathy. Naunyn Schmiedeberg Arch Pharmacol. 2021;394(6):1301–14. 10.1007/s00210-020-02040-8
14. Abdoli N, Sadeghian I, Azarpira N, Ommati MM, Heidari R. Taurine mitigates bile duct obstruction-associated cholemic nephropathy: Effect on oxidative stress and mitochondrial parameters. Clin Exp Hepatol. 2021;7(1):30–40. 10.5114/ceh.2021.104675
15. Jamshidzadeh A, Heidari R, Golzar T, Derakhshanfar A. Effect of Eisenia foetida extract against cisplatin-induced kidney injury in rats. J Diet Suppl. 2016;13(5):551–9. 10.3109/19390211.2015.1124163
16. Jamshidzadeh A, Heidari R, Mohammadi-Samani S, Azarpira N, Najbi A, Jahani P, et al. A comparison between the nephrotoxic profile of gentamicin and gentamicin nanoparticles in mice. J Biochem Mol Toxicol. 2015;29(2):57–62. 10.1002/jbt.21667
17. Gyurászová M, Gurecká R, Bábícˇková J, Tóthová L’. Oxidative stress in the pathophysiology of kidney disease: Implications for noninvasive monitoring and identification of biomarkers. Oxid Med Cell Longev. 2020;2020:5478708. 10.1155/2020/5478708
18. Pavlakou P, Liakopoulos V, Eleftheriadis T, Mitsis M, Dounousi E. Oxidative stress and acute kidney injury in critical illness: Pathophysiologic mechanisms-biomarkers-interventions, and future perspectives. Oxid Med Cell Longev. 2017;2017:e6193694. 10.1155/2017/6193694
19. Zhao Y-H, Shen C-F, Wang G-J, Kang Y, Song Y-H, Liu J-W. Curcumin alleviates acute kidney injury in a dry-heat environment by reducing oxidative stress and inflammation in a rat model. J Biochem Mol Toxicol. 2021;35(1):e22630. 10.1002/jbt.22630
20. Brookes PS, Yoon Y, Robotham JL, Anders MW, Sheu S-S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am J Physiol. 2004;287(4):C817–33. 10.1152/ajpcell.00139.2004
21. Heidari R, Ghanbarinejad V, Mohammadi H, Ahmadi A, Ommati MM, Abdoli N, et al. Mitochondria protection as a mechanism underlying the hepatoprotective effects of glycine in cholestatic mice. Biomed Pharmacother. 2018;97:1086–95. 10.1016/j.biopha.2017.10.166
22. Khodaei F, Rashedinia M, Heidari R, Rezaei M, Khoshnoud MJ. Ellagic acid improves muscle dysfunction in cuprizone-induced demyelinated mice via mitochondrial Sirt3 regulation. Life Sci. 2019;237:116954. 10.1016/j.lfs.2019.116954
23. Jamshidzadeh A, Niknahad H, Heidari R, Azadbakht M, Khodaei F, Arabnezhad MR, Farshad O. Propylthiouracil-induced mitochondrial dysfunction in liver and its relevance to drug-induced hepatotoxicity. Pharm Sci. 2017;23(2):95–102. 10.15171/PS.2017.15
24. Ghanbarinejad V, Ommati MM, Jia Z, Farshad O, Jamshidzadeh A, Heidari R. Disturbed mitochondrial redox state and tissue energy charge in cholestasis. J Biochem Mol Toxicol. 2021;35(9):e22846. 10.1002/jbt.22846
25. Heidari R, Ommati MM, Niknahad H. Mitochondria as biosynthetic centers and targeted therapeutics. In: Pourahmad J, Rezaei M, editors. Mitochondrial metabolism. Cambridge, MA: Academic Press; 2021, pp. 19–47.
26. Heidari R, Ommati MM, Niknahad H. Ammonia. In: de Oliveira MR, editor. Mitochondrial intoxication. Cambridge, MA: Academic Press; 2023, pp. 249–70.
27. Ommati MM, Mohammadi H, Mousavi K, Azarpira N, Farshad O, Dehghani R, et al. Metformin alleviates cholestasis-associated nephropathy through regulating oxidative stress and mitochondrial function. Liver Res. 2021;5(3):171–80. 10.1016/j.livres.2020.12.001
28. Mousavi K, Niknahad H, Ghalamfarsa A, Mohammadi H, Azarpira N, Ommati MM, et al. Taurine mitigates cirrhosis-associated heart injury through mitochondrial-dependent and antioxidative mechanisms. Clin Exp Hepatol. 2020;6(3):207–19. 10.5114/ceh.2020.99513
29. Heidari R. The footprints of mitochondrial impairment and cellular energy crisis in the pathogenesis of xenobiotics-induced nephrotoxicity, serum electrolytes imbalance, and Fanconi’s syndrome: A comprehensive review. Toxicology. 2019;423:1–31. 10.1016/j.tox.2019.05.002
30. Heidari R, Ommati MM, Niknahad H. Drug-induced mitochondrial impairment: Mechanisms and testing systems. In: de Oliveira MR, editor. Mitochondrial intoxication. Cambridge, MA: Academic Press; 2023, pp. 49–76.
31. Tang C, Dong Z. Mitochondria in kidney injury: When the power plant fails. J Am Soc Nephrol. 2016;27(7):1869–72. 10.1681/ASN.2015111277
32. Niknahad AM, Ommati MM, Farshad O, Moezi L, Heidari R. Manganese-induced nephrotoxicity is mediated through oxidative stress and mitochondrial impairment. J Renal Hepatic Disorder. 2020;4(2):1–10. 10.15586/jrenhep.2020.66
33. Chevalier RL. Pathogenesis of renal injury in obstructive uropathy. Curr Opin Pediatr. 2006;18(2):153–60. 10.1097/01.mop.0000193287.56528.a4
34. Chávez-Iñiguez JS, Navarro-Gallardo GJ, Medina-González R, Alcantar-Vallin L, García-García G. Acute kidney injury caused by obstructive nephropathy. Int J Nephrol. 2020. eCollection 2020: 8846622. 10.1155/2020/8846622
35. Hesketh EE, Vernon MA, Ding P, Clay S, Borthwick G, Conway B, et al. A murine model of irreversible and reversible unilateral ureteric obstruction. J Vis Exp. 201494:e52559. 10.3791/52559
36. Heidari R, Mohammadi H, Ghanbarinejad V, Ahmadi A, Ommati MM, Niknahad H, et al. Proline supplementation mitigates the early stage of liver injury in bile duct ligated rats. J Basic Clin Physiol Pharmacol. 2019;30(1):91–101. 10.1515/jbcpp-2017-0221
37. Heidari R, Ghanbarinejad V, Mohammadi H, Ahmadi A, Esfandiari A, Azarpira N, et al. Dithiothreitol supplementation mitigates hepatic and renal injury in bile duct ligated mice: Potential application in the treatment of cholestasis-associated complications. Biomed Pharmacother. 2018;99:1022–32. 10.1016/j.biopha.2018.01.018
38. Heidari R, Arabnezhad MR, Ommati MM, Azarpira N, Ghodsimanesh E, Niknahad H. Boldine supplementation regulates mitochondrial function and oxidative stress in a rat model of hepatotoxicity. Pharm Sci. 2019;25(1):1–10. 10.15171/PS.2019.1
39. Ommati MM, Sabouri S, Niknahad H, Arjmand A, Alidaee S, Mazloomi S, et al. Pulmonary inflammation, oxidative stress, and fibrosis in a mouse model of cholestasis: The potential protective properties of the dipeptide carnosine. Naunyn Schmiedeberg Arch Pharmacol. 2023;396:1129–42. 10.1007/s00210-023-02391-y
40. Ommati MM, Sabouri S, Retana-Marquez S, Nategh Ahmadi H, Arjmand A, Alidaee S, et al. Taurine improves sperm mitochondrial indices, blunts oxidative stress parameters, and enhances steroidogenesis and kinematics of sperm in lead-exposed mice. Reprod Sci. 2023;30(6):1891–910. 10.1007/s43032-022-01140-5
41. Ommati MM, Niknahad H, Najibi A, Arjmand A, Alidaee S, Mazloomi S, et al. Cholestasis-associated pulmonary inflammation, oxidative stress, and tissue fibrosis: The protective role of the biogenic amine agmatine. Pharmacology. 2023:378–92. 10.1159/000530307
42. Heidari R, Jamshidzadeh A, Ghanbarinejad V, Ommati MM, Niknahad H. Taurine supplementation abates cirrhosis-associated locomotor dysfunction. Clin Exp Hepatol. 2018;4(2):72–82. 10.5114/ceh.2018.75956
43. Heidari R, Behnamrad S, Khodami Z, Ommati MM, Azarpira N, Vazin A. The nephroprotective properties of taurine in colistin-treated mice is mediated through the regulation of mitochondrial function and mitigation of oxidative stress. Biomed Pharmacother. 2019;109:103–11. 10.1016/j.biopha.2018.10.093
44. Niknahad H, Heidari R, Alzuhairi AM, Najibi A. Mitochondrial dysfunction as a mechanism for pioglitazone-induced injury toward HepG2 cell line. Pharm Sci. 2015;20(4):169–74. 10.5681/PS.2015.007
45. Ommati MM, Amjadinia A, Mousavi K, Azarpira N, Jamshidzadeh A, Heidari R. N-acetyl cysteine treatment mitigates biomarkers of oxidative stress in different tissues of bile duct ligated rats. Stress. 2021;24(2):213–28. 10.1080/10253890.2020.1777970
46. Owji H, Hemmati S, Heidari R, Hakimzadeh M. Effect of alumina (Al2O3) nanoparticles and macroparticles on Trigonella foenum-graceum L. in vitro cultures: Assessment of growth parameters and oxidative stress-related responses. 3 Biotech. 2019;9(11):419. 10.1007/s13205-019-1954-7
47. Gholami A, Montazeri-Najafabady N, Ashoori Y, Kazemi K, Heidari R, Omidifar N, et al. The ameliorating effect of limosilactobacillus fermentum and its supernatant post-biotic on cisplatin-induced chronic kidney disease in an animal model. BMC Complement Med Therap. 2023;23(1):243. 10.1186/s12906-023-04068-8
48. Ommati MM, Mobasheri A, Ma Y, Xu D, Tang Z, Manthari RK, et al. Taurine mitigates the development of pulmonary inflammation, oxidative stress, and histopathological alterations in a rat model of bile duct ligation. Naunyn Schmiedeberg Arch Pharmacol. 2022;395(12):1557–72. 10.1007/s00210-022-02291-7
49. Eftekhari A, Ahmadian E, Azami A, Johari-Ahar M, Eghbal MA. Protective effects of coenzyme Q10 nanoparticles on dichlorvos-induced hepatotoxicity and mitochondrial/lysosomal injury. Environ Toxicol. 2018;33(2):167–77. 10.1002/tox.22505
50. Zhang Y-T, Zheng Q-S, Pan J, Zheng R-L. Oxidative damage of biomolecules in mouse liver induced by morphine and protected by antioxidants. Basic Clin Pharmacol Toxicol. 2004;95(2): 53–8. 10.1111/j.1742-7843.2004.950202.x
51. Heidari R, Babaei H, Eghbal MA. Amodiaquine-induced toxicity in isolated rat hepatocytes and the cytoprotective effects of taurine and/or N-acetyl cysteine. Res Pharm Sci. 2014;9(2):97–105.
52. Ommati MM, Ahmadi HN, Sabouri S, Retana-Marquez S, Abdoli N, Rashno S, et al. Glycine protects the male reproductive system against lead toxicity via alleviating oxidative stress, preventing sperm mitochondrial impairment, improving kinematics of sperm, and blunting the downregulation of enzymes involved in the steroidogenesis. Environ Toxicol. 2022;37(12):2990–3006. 10.1002/tox.23654
53. Ommati MM, Farshad O, Niknahad H, Arabnezhad MR, Azarpira N, Mohammadi HR, et al. Cholestasis-associated reproductive toxicity in male and female rats: The fundamental role of mitochondrial impairment and oxidative stress. Toxicol Lett. 2019;316:60–72. 10.1016/j.toxlet.2019.09.009
54. Niknahad H, Heidari R, Firuzi R, Abazari F, Ramezani M, Azarpira N, et al. Concurrent inflammation augments antimalarial drugs-induced liver injury in rats. Adv Pharm Bull. 2016;6(4):617–25. 10.15171/apb.2016.076
55. Farshad O, Heidari R, Zamiri MJ, Retana-Márquez S, Khalili M, Ebrahimi M, et al. Spermatotoxic effects of single-walled and multi-walled carbon nanotubes on male mice. Front Vet Sci. 2020;7:591558. 10.3389/fvets.2020.591558
56. Abdoli N, Sadeghian I, Mousavi K, Azarpira N, Ommati MM, Heidari R. Suppression of cirrhosis-related renal injury by N-acetyl cysteine. Curr Res Pharmacol Drug Discov. 2020;1:30–8. 10.1016/j.crphar.2020.100006
57. Heidari R, Babaei H, Eghbal MA. Cytoprotective effects of organosulfur compounds against methimazole induced toxicity in isolated rat hepatocytes. Adv Pharm Bull. 2013;3(1):135–42. 10.5681/apb.2013.023
58. Heidari R, Moezi L, Asadi B, Ommati MM, Azarpira N. Hepatoprotective effect of boldine in a bile duct ligated rat model of cholestasis/cirrhosis. PharmaNutrition. 2017;5(3):109–17. 10.1016/j.phanu.2017.07.001
59. Ommati MM, Abdoli N, Firoozi M, Akhlagh A, Mazloomi S, Mousavi K, et al. Sildenafil blunts lung inflammation and oxidative stress in a rat model of cholestasis. Pharm Sci. 2023;29(2):165–74. 10.34172/PS.2022.38
60. Pasdaran A, Azarpira N, Heidari R, Nourinejad S, Zare M, Hamedi A. Effects of some cosmetic dyes and pigments on the proliferation of human foreskin fibroblasts and cellular oxidative stress; potential cytotoxicity of chlorophyllin and indigo carmine on fibroblasts. J Cosmet Dermatol. 2022;21(9):3979–85. 10.1111/jocd.14695
61. Niknahad H, Nadgaran A, Alidaee S, Arjmand A, Abdoli N, Mazloomi SM, et al. Thiol-reducing agents abate cholestasis-induced lung inflammation, oxidative stress, and histopathological alterations. Trend Pharm Sci. 2023;9(1):55–70. 10.30476/tips.2023.96536.1165
62. Heidari R, Ommati MM, Alahyari S, Azarpira N, Niknahad H. Amino acid-containing krebs-henseleit buffer protects rat liver in a long-term organ perfusion model. Pharm Sci. 2018;24(3):168–79. 10.15171/PS.2018.25
63. Heidari R, Niknahad H, Jamshidzadeh A, Azarpira N, Bazyari M, Najibi A. Carbonyl traps as potential protective agents against methimazole-induced liver injury. J Biochem Mol Toxicol. 2015;29(4):173–81. 10.1002/jbt.21682
64. Heidari R, Esmailie N, Azarpira N, Najibi A, Niknahad H. Effect of thiol-reducing agents and antioxidants on sulfasalazine-induced hepatic injury in normotermic recirculating isolated perfused rat liver. Toxicol Res. 2016;32(2):133–40. 10.5487/TR.2016.32.2.133
65. Najibi A, Rezaei H, Manthari RK, Niknahad H, Jamshidzadeh A, Farshad O, et al. Cellular and mitochondrial taurine depletion in bile duct ligated rats: A justification for taurine supplementation in cholestasis/cirrhosis. Clin Exp Hepatol. 2022;8(3):195–210. 10.5114/ceh.2022.119216
66. Niknahad H, Mehrabani PS, Arjmand A, Alidaee S, Mazloomi S, Ahmadi P, et al. Cirrhosis-induced oxidative stress in erythrocytes: The therapeutic potential of taurine. Clin Exp Hepatol. 2023;9(1):79–92. 10.5114/ceh.2023.126028
67. Ommati MM, Mobasheri A, Niknahad H, Rezaei M, Alidaee S, Arjmand A, et al. Low-dose ketamine improves animals’ locomotor activity and decreases brain oxidative stress and inflammation in ammonia-induced neurotoxicity. J Biochem Mol Toxicol. 2023:e23468. 10.1002/jbt.23468
68. Niknahad H, Abdoli N, Ommati MM, Najibi A, Zare A, Mazloomi S, et al. Dexamethasone blunts lung inflammation in cholestatic mice. Trend Pharm Sci. 2023;9(1):1–14. 10.30476/tips.2022.96046.1155
69. Ommati MM, Arabnezhad MR, Farshad O, Jamshidzadeh A, Niknahad H, Retana-Marquez S, et al. The role of mitochondrial impairment and oxidative stress in the pathogenesis of lithium-induced reproductive toxicity in male mice. Front Vet Sci. 2021;8:603262. 10.3389/fvets.2021.603262
70. Karamikhah R, Jamshidzadeh A, Azarpira N, Saeidi A, Heidari R. Propylthiouracil-induced liver injury in mice and the protective role of taurine. Pharm Sci. 2016;21(2):94–101. 10.15171/PS.2015.23
71. Niknahad H, Heidari R, Mohammadzadeh R, Ommati MM, Khodaei F, Azarpira N, et al. Sulfasalazine induces mitochondrial dysfunction and renal injury. Ren Fail. 2017;39(1):745–53. 10.1080/0886022X.2017.1399908
72. Ommati MM, Jamshidzadeh A, Saeed M, Rezaei M, Heidari R. Dextromethorphan improves locomotor activity and decreases brain oxidative stress and inflammation in an animal model of acute liver failure. Clin Exp Hepatol. 2022;8(3):178–87. 10.5114/ceh.2022.118299
73. Mobasher P, Ommati MM, Najibi A, Akbarizadeh AR, Ghasemi Y, Heidari R. Searching for alternative toxicology testing systems: The response of isolated mitochondria from Saccharomyces cerevisiae, potato tuber, and mouse liver to a toxic insult. Trend Pharm Sci. 2021;7(3):179–90. 10.30476/tips.2021.91544.1102
74. Ahmadi N, Rezaee Z, Azarpira N, Zahedi S, Saeedi A, Jamshidzadeh A, et al. A histopathological evaluation on the effect of captopril in cyclophosphamide-induced hemorrhagic cystitis. Trend Pharm Sci. 2021;7(1):35–48. 10.30476/tips.2021.90418.1085
75. Hamdy N, El-Demerdash E. New therapeutic aspect for carvedilol: Antifibrotic effects of carvedilol in chronic carbon tetrachloride-induced liver damage. Toxicol Appl Pharmacol. 2012;261(3):292–9. 10.1016/j.taap.2012.04.012
76. Jamshidzadeh A, Heidari R, Latifpour Z, Ommati MM, Abdoli N, Mousavi S, et al. Carnosine ameliorates liver fibrosis and hyperammonemia in cirrhotic rats. Clin Res Hepatol Gastroenterol. 2017;41(4):424–34. 10.1016/j.clinre.2016.12.010
77. Ommati MM, Jamshidzadeh A, Heidari R, Sun Z, Zamiri MJ, Khodaei F, et al. Carnosine and histidine supplementation blunt lead-induced reproductive toxicity through antioxidative and mitochondria-dependent mechanisms. Biol Trace Elem Res. 2019;187(1):151–62. 10.1007/s12011-018-1358-2
78. Niknahad H, Ommati MM, Sookhak N, Hajihashemi F, Azarpira N, Heidari R. Glycyrrhizic acid and the aqueous extract of glycyrrhiza glabra attenuate hepatotoxicity in mice. Trend Pharm Sci. 2021;7(1):59–72. 10.30476/tips.2021.90843.1090
79. Ommati MM, Hojatnezhad S, Abdoli N, Manthari RK, Jia Z, Najibi A, et al. Pentoxifylline mitigates cholestasis-related cholemic nephropathy. Clin Exp Hepatol. 2021;7(4):377–89. 10.5114/ceh.2021.111014
80. Ommati MM, Farshad O, Niknahad H, Mousavi K, Moein M, Azarpira N, et al. Oral administration of thiol-reducing agents mitigates gut barrier disintegrity and bacterial lipopolysaccharide translocation in a rat model of biliary obstruction. Curr Res Pharm Drug Discov. 2020;1:10–8. 10.1016/j.crphar.2020.06.001
81. Heidari R, Ahmadi A, Ommati MM, Niknahad H. Methylene blue improves mitochondrial function in the liver of cholestatic rats. Trend Pharm Sci. 2020;6(2):73–86. 10.30476/tips.2020.85961.1043
82. Akram J, Reza H, Farzaneh A, Maral R, Forouzan K, Mohammad Mehdi O, et al. Antimalarial drugs-induced hepatic injury in rats and the protective role of carnosine. Pharm Sci. 2016;22(3):170–80. 10.15171/PS.2016.27
83. Ommati MM, Farshad O, Ghanbarinejad V, Mohammadi HR, Khadijeh M, Negar A, et al. The nephroprotective role of carnosine against ifosfamide-induced renal injury and electrolytes imbalance is mediated via the regulation of mitochondrial function and alleviation of oxidative stress. Drug Res. 2020;70(1):49–56. 10.1055/a-1017-5085
84. Liu SF, Ye X, Malik AB. Pyrrolidine dithiocarbamate prevents I-κB degradation and reduces microvascular injury induced by lipopolysaccharide in multiple organs. Mol Pharmacol. 1999;55(4):658–67.
85. Ommati MM, Manthari RK, Tikka C, Niu R, Sun Z, Sabouri S, et al. Arsenic-induced autophagic alterations and mitochondrial impairments in HPG-S axis of mature male mice offspring (F1-generation): A persistent toxicity study. Toxicol Lett. 2020;326:83–98. 10.1016/j.toxlet.2020.02.013
86. Ommati MM, Shi X, Li H, Zamiri MJ, Farshad O, Jamshidzadeh A, et al. The mechanisms of arsenic-induced ovotoxicity, ultrastructural alterations, and autophagic related paths: An enduring developmental study in folliculogenesis of mice. Ecotoxicol Environ Saf. 2020;204:110973. 10.1016/j.ecoenv.2020.110973
87. Farshad O, Ommati MM, Yüzügülen J, Jamshidzadeh A, Mousavi K, Ahmadi Z, et al. Carnosine mitigates biomarkers of oxidative stress, improves mitochondrial function, and alleviates histopathological alterations in the renal tissue of cholestatic rats. Pharm Sci. 2020;27(1):32–45. 10.34172/PS.2020.60
88. Mousavi K, Manthari RK, Najibi A, Jia Z, Ommati MM, Heidari R. Mitochondrial dysfunction and oxidative stress are involved in the mechanism of tramadol-induced renal injury. Curr Res Pharmacol Drug Discov. 2021;2:100049. 10.1016/j.crphar.2021.100049
89. Niknahad H, Jamshidzadeh A, Heidari R, Abdoli N, Ommati MM, Jafari F, et al. The postulated hepatotoxic metabolite of methimazole causes mitochondrial dysfunction and energy metabolism disturbances in liver. Pharm Sci. 2016;22(4):217–26. 10.15171/PS.2016.35
90. Caro AA, Adlong LW, Crocker SJ, Gardner MW, Luikart EF, Gron LU. Effect of garlic-derived organosulfur compounds on mitochondrial function and integrity in isolated mouse liver mitochondria. Toxicol Lett. 2012;214(2):166–74. 10.1016/j.toxlet.2012.08.017
91. Ahmadi N, Ghanbarinejad V, Ommati MM, Jamshidzadeh A, Heidari R. Taurine prevents mitochondrial membrane permeabilization and swelling upon interaction with manganese: Implication in the treatment of cirrhosis-associated central nervous system complications. J Biochem Mol Toxicol. 2018;32(11):e22216. 10.1002/jbt.22216
92. Ahmadian E, Khosroushahi AY, Eghbal MA, Eftekhari A. Betanin reduces organophosphate induced cytotoxicity in primary hepatocyte via an anti-oxidative and mitochondrial dependent pathway. Pestic Biochem Physiol. 2018;144:71–8. 10.1016/j.pestbp.2017.11.009
93. Khodaei F, Khoshnoud MJ, Heidaryfar S, Heidari R, Karimpour Baseri MH, Azarpira N, et al. The effect of ellagic acid on spinal cord and sciatica function in a mice model of multiple sclerosis. J Biochem Mol Toxicol. 2020;34(11):e22564. 10.1002/jbt.22564
94. Heidari R, Rasti M, Shirazi Yeganeh B, Niknahad H, Saeedi A, Najibi A. Sulfasalazine-induced renal and hepatic injury in rats and the protective role of taurine. BioImpacts. 2016;6(1):3–8. 10.15171/bi.2016.01
95. Ommati MM, Heidari R, Manthari RK, Tikka Chiranjeevi S, Niu R, Sun Z, et al. Paternal exposure to arsenic resulted in oxidative stress, autophagy, and mitochondrial impairments in the HPG axis of pubertal male offspring. Chemosphere. 2019;236:124325. 10.1016/j.chemosphere.2019.07.056
96. Ommati MM, Azarpira N, Khodaei F, Niknahad H, Gozashtegan V, Heidari R. Methylene blue treatment enhances mitochondrial function and locomotor activity in a C57BL/6 mouse model of multiple sclerosis. Trend Pharm Sci. 2020;6(1):29–42. 10.30476/tips.2020.85962.1044
97. Eftekhari A, Azarmi Y, Parvizpur A, Eghbal MA. Involvement of oxidative stress and mitochondrial/lysosomal cross-talk in olanzapine cytotoxicity in freshly isolated rat hepatocytes. Xenobiotica. 2016;46(4):369–78. 10.3109/00498254.2015.1078522
98. Ommati MM, Heidari R, Ghanbarinejad V, Abdoli N, Niknahad H. Taurine treatment provides neuroprotection in a mouse model of manganism. Biol Trace Elem Res. 2019;190(2):384–95. 10.1007/s12011-018-1552-2
99. Eftekhari A, Heidari R, Ahmadian E, Eghbal MA. Cytoprotective properties of carnosine against isoniazid-induced toxicity in primary cultured rat hepatocytes. Pharm Sci. 2018;24(4):257–63. 10.15171/PS.2018.38
100. Heidari R, Jamshidzadeh A, Ommati MM, Rashidi E, Khodaei F, Sadeghi A, et al. Ammonia-induced mitochondrial impairment is intensified by manganese co-exposure: relevance to the management of subclinical hepatic encephalopathy and cirrhosis-associated brain injury. Clin Exp Hepatol. 2019;5(2):109–17. 10.5114/ceh.2019.85071
101. Ommati MM, Farshad O, Mousavi K, Taghavi R, Farajvajari S, Azarpira N, et al. Agmatine alleviates hepatic and renal injury in a rat model of obstructive jaundice. PharmaNutrition. 2020;13:100212. 10.1016/j.phanu.2020.100212
102. Jamshidzadeh A, Niknahad H, Heidari R, Zarei M, Ommati MM, Khodaei F. Carnosine protects brain mitochondria under hyperammonemic conditions: Relevance to hepatic encephalopathy treatment. PharmaNutrition. 2017;5(2):58–63. 10.1016/j.phanu.2017.02.004
103. Heidari R, Mousavi K, Amin S, Ommati MM, Niknahad H. N-acetylcysteine treatment protects intestinal mitochondria in a surgical stress model. Trend Pharm Sci. 2020;6(2):87–96. 10.30476/tips.2020.85960.1042
104. Seifi K, Rezaei M, Yansari AT, Riazi GH, Zamiri MJ, Heidari R. Saturated fatty acids may ameliorate environmental heat stress in broiler birds by affecting mitochondrial energetics and related genes. J Therm Biol. 2018;78:1–9. 10.1016/j.jtherbio.2018.08.018
105. Niknahad H, Jamshidzadeh A, Heidari R, Hosseini Z, Mobini K, Khodaei F, et al. Paradoxical effect of methimazole on liver mitochondria: In vitro and in vivo. Toxicol Lett. 2016;259:108–15. 10.1016/j.toxlet.2016.08.003
106. Ommati MM, Farshad O, Mousavi K, Jamshidzadeh A, Azmoon M, Heidari S, et al. Betaine supplementation mitigates intestinal damage and decreases serum bacterial endotoxin in cirrhotic rats. PharmaNutrition. 2020;12:100179. 10.1016/j.phanu.2020.100179
107. Iravanpour F, Dargahi L, Rezaei M, Haghani M, Heidari R, Valian N, et al. Intranasal insulin improves mitochondrial function and attenuates motor deficits in a rat 6-OHDA model of Parkinson’s disease. CNS Neurosci Ther. 2021;27(3):308–19. 10.1111/cns.13609
108. Heidari R, Abdoli N, Ommati MM, Jamshidzadeh A, Niknahad H. Mitochondrial impairment induced by chenodeoxycholic acid: The protective effect of taurine and carnosine supplementation. Trend Pharm Sci. 2018;4(2):99–108. 10.1111/tips.v4i2.182
109. Ghanbarinejad V, Ahmadi A, Niknahad H, Ommati MM, Heidari R. Carnosine mitigates manganese mitotoxicity in an in vitro model of isolated brain mitochondria. Adv Pharm Bull. 2019;9(2):294–301. 10.15171/apb.2019.034
110. Ommati MM, Farshad O, Jamshidzadeh A, Heidari R. Taurine enhances skeletal muscle mitochondrial function in a rat model of resistance training. PharmaNutrition. 2019;9:100161. 10.1016/j.phanu.2019.100161
111. Mohammadi H, Sayad A, Mohammadi M, Niknahad H, Heidari R. N-acetyl cysteine treatment preserves mitochondrial indices of functionality in the brain of hyperammonemic mice. Clin Exp Hepatol. 2020;6(2):106–15. 10.5114/ceh.2020.95814
112. Ommati MM, Farshad O, Mousavi K, Khalili M, Jamshidzadeh A, Heidari R. Chlorogenic acid supplementation improves skeletal muscle mitochondrial function in a rat model of resistance training. Biologia. 2020;75(8):1221–30. 10.2478/s11756-020-00429-7
113. Farshad O, Ommati MM, Yüzügülen J, Alizadeh S, Mousavi K, Azarpira N, et al. Skeletal muscle mitochondrial impairment in cirrhosis-induced sarcopenia. Trend Pharm Sci. 2020;6(3):189–204. 10.30476/tips.2020.87789.1067
114. Heidari R, Ghanbarinejad V, Ommati MM, Jamshidzadeh A, Niknahad H. Mitochondria protecting amino acids: Application against a wide range of mitochondria-linked complications. PharmaNutrition. 2018;6(4):180–90. 10.1016/j.phanu.2018.09.001
115. Heidari R, Mandegani L, Ghanbarinejad V, Siavashpour A, Ommati MM, Azarpira N, et al. Mitochondrial dysfunction as a mechanism involved in the pathogenesis of cirrhosis-associated cholemic nephropathy. Biomed Pharmacother. 2019;109:271–80. 10.1016/j.biopha.2018.10.104
116. Truong DH, Eghbal MA, Hindmarsh W, Roth SH, O’Brien PJ. Molecular mechanisms of hydrogen sulfide toxicity. Drug Metab Rev. 2006;38(4):733–44. 10.1080/03602530600959607
117. Najafi N, Jamshidzadeh A, Fallahzadeh H, Omidi M, Abdoli N, Najibi A, et al. Valproic acid-induced hepatotoxicity and the protective role of thiol reductants. Trends Pharm Sci. 2017;3(2):63–70.
118. Ommati MM, Li H, Jamshidzadeh A, Khoshghadam F, Retana-Márquez S, Lu Y, et al. The crucial role of oxidative stress in non-alcoholic fatty liver disease-induced male reproductive toxicity: The ameliorative effects of Iranian indigenous probiotics. Naunyn Schmiedeberg Arch Pharmacol. 2022:1–19. 10.1007/s00210-021-02177-0
119. Niknahad H, Jamshidzadeh A, Heidari R, Zarei M, Ommati MM. Ammonia-induced mitochondrial dysfunction and energy metabolism disturbances in isolated brain and liver mitochondria, and the effect of taurine administration: relevance to hepatic encephalopathy treatment. Clin Exp Hepatol. 2017;3(3):141–51. 10.5114/ceh.2017.68833
120. Siavashpour A, Khalvati B, Azarpira N, Mohammadi H, Niknahad H, Heidari R. Poly (ADP-ribose) polymerase-1 (PARP-1) overactivity plays a pathogenic role in bile acids-induced nephrotoxicity in cholestatic rats. Toxicol Lett. 2020;330:144–58. 10.1016/j.toxlet.2020.05.012
121. Ommati MM, Heidari R, Ghanbarinejad V, Aminian A, Abdoli N, Niknahad H. The neuroprotective properties of carnosine in a mouse model of manganism is mediated via mitochondria regulating and antioxidative mechanisms. Nutr Neurosci. 2020;23(9):731–43. 10.1080/1028415X.2018.1552399
122. Heidari R, Taheri V, Rahimi HR, Shirazi Yeganeh B, Niknahad H, Najibi A. Sulfasalazine-induced renal injury in rats and the protective role of thiol-reductants. Ren Fail. 2016;38(1):137–41. 10.3109/0886022X.2015.1096731
123. Li W, Lu Y, Lou Y, Zhao S, Cui W, Wang Y, et al. FFNT25 ameliorates unilateral ureteral obstruction-induced renal fibrosis. Ren Fail. 2019;41(1):419–26. 10.1080/0886022X.2019.1612430
124. Riyach O, Ahsaini M, Kharbach Y, Bounoual M, Tazi MF, El Ammari JE, et al. Bilateral ureteral obstruction revealing a benign prostatic hypertrophy: A case report and review of the literature. J Med Case Rep. 2014;8:42. 10.1186/1752-1947-8-42
125. Jung JH, Park J, Kim WT, Kim HW, Kim HJ, Hong S, et al. The association of benign prostatic hyperplasia with lower urinary tract stones in adult men: A retrospective multicenter study. Asian J Urol. 2018;5(2):118–21. 10.1016/j.ajur.2017.06.008
126. Kawada N, Moriyama T, Ando A, Fukunaga M, Miyata T, Kurokawa K, et al. Increased oxidative stress in mouse kidneys with unilateral ureteral obstruction. Kidney Int. 1999;56(3):1004–13. 10.1046/j.1523-1755.1999.00612.x
127. Martínez-Klimova E, Aparicio-Trejo OE, Tapia E, Pedraza-Chaverri J. Unilateral ureteral obstruction as a model to investigate fibrosis-attenuating treatments. Biomolecules. 2019;9(4):141. 10.3390/biom9040141
128. Li Z, Li A, Gao J, Li H, Qin X. Kidney tissue targeted metabolic profiling of unilateral ureteral obstruction rats by NMR. Front Pharmacol. 2016;7:307. 10.3389/fphar.2016.00307
129. Lee W-C, Jao H-Y, Hsu J-D, Lee Y-R, Wu M-J, Kao Y-L, et al. Apple polyphenols reduce inflammation response of the kidneys in unilateral ureteral obstruction rats. J Funct Foods. 2014;11:1–11. 10.1016/j.jff.2014.08.010
130. Schulze-Osthoff K, Bakker AC, Vanhaesebroeck B, Beyaert R, Jacob WA, Fiers W. Cytotoxic activity of tumor necrosis factor is mediated by early damage of mitochondrial functions. Evidence for the involvement of mitochondrial radical generation. J Biol Chem. 1992;267(8):5317–23. 10.1016/S0021-9258(18)42768-8
131. Shen Y-L, Shi Y-Z, Chen G-G, Wang L-L, Zheng M-Z, Jin H-F, et al. TNF-α induces Drp1-mediated mitochondrial fragmentation during inflammatory cardiomyocyte injury. Int J Mol Med. 2018;41(4):2317–27. 10.3892/ijmm.2018.3385
132. Mahmoud AM, Wilkinson FL, Sandhu MA, Lightfoot AP. The interplay of oxidative stress and inflammation: Mechanistic insights and therapeutic potential of antioxidants. Oxidative Med Cell Long. 2021;2021:1–4. 10.1155/2021/9851914
133. Donia T, Khamis A. Management of oxidative stress and inflammation in cardiovascular diseases: Mechanisms and challenges. Envir Sci Poll Res. 2021;28(26):34121–53. 10.1007/s11356-021-14109-9
134. Emadi E, Abdoli N, Ghanbarinejad V, Mohammadi HR, Mousavi Mobarakeh K, Azarpira N, et al. The potential role of mitochondrial impairment in the pathogenesis of imatinib-induced renal injury. Heliyon. 2019;5(6):e01996. 10.1016/j.heliyon.2019.e01996
135. Heidari R. Brain mitochondria as potential therapeutic targets for managing hepatic encephalopathy. Life Sci. 2019;218:65–80. 10.1016/j.lfs.2018.12.030
136. Tait SWG, Green DR. Mitochondrial regulation of cell death. Cold Spring Harb Perspect Biol. 2013;5(9): a008706. 10.1101/cshperspect.a008706
137. Mohammadi H, Ommati MM, Farshad O, Jamshidzadeh A, Nikbakht MR, Niknahad H, et al. Taurine and isolated mitochondria: A concentration-response study. Trend Pharm Sci. 2019;5(4):197–206. 10.30476/tips.2020.84851.1037
138. Vazin A, Heidari R, Khodami Z. Curcumin supplementation alleviates polymyxin E-induced nephrotoxicity. J Exp Pharmacol. 2020;12:129–36. 10.2147/JEP.S255861
139. Ommati MM, Heidari R. Amino acids ameliorate heavy metals-induced oxidative stress in male/female reproductive tissue. In: Toxicology. Editors: Vinood B. Patel and Victor R. Preedy New York, NY: Elsevier; 2021, pp. 371–86.